日本基础生物学研究所植物环境应答研究部门的森田(寺尾)美代教授、西村岳志助教和中村守贵特任研究员、福建农林大学的古谷将彦教授以及东京大学的平野良宪助教,与奈良先端科学技术大学院大学的箱嶋敏雄教授、基础生物学研究所生命创成探究中心的加藤辉特任助教以及立命馆大学的深尾阳一郎副教授等联合研究,利用模式植物拟南芥发现了重力感受细胞中与重力方向的信号传递有关的新因子RLD。
植物的根部朝着地下、茎部朝着空中生长。这是植物感测重力(地心引力)方向发生的向地性(向重力性)反应。此前虽然知道重力的方向通过根和茎的特殊细胞——重力感受细胞感知,植物向性通过控制生长素的传输实现对重力方向产生反应等,但一直不清楚重力感受细胞内的重力方向信息传递机制的详细情况。森田教授等人组成的研究团队此前确认,名为LZY的蛋白质是向重力性所需的物质。此次新发现了与LZY相互作用的蛋白质RLD,同时确认,RLD也是与向重力性有关的重要因子,RLD参与了生长素的传输调控。并提出生长素的传输调控分子机制,即LZY响应重力方向改变其在重力感受细胞内的位置,此时结合RLD并将其置于同一位置。相关研究成果已于2020年1月3日发布在国际学术期刊《Nature Communications》上。
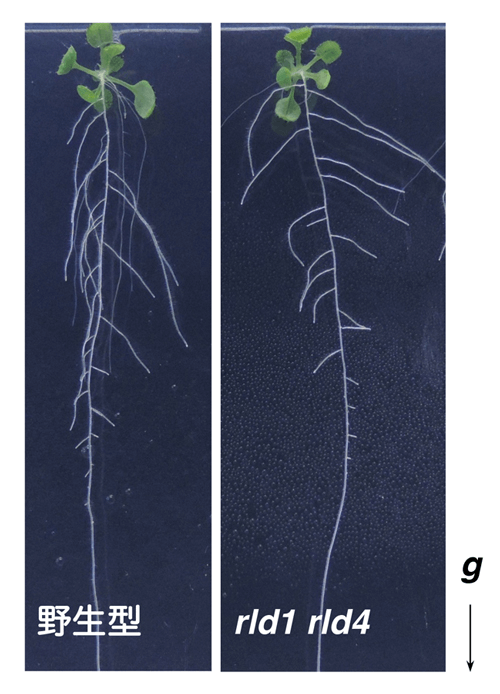
图:通常,植物根部会感测重力的方向,表现出沿重力方向生长的向重力性(左:野生型);但在rld1 rld4双重突变体(右)中,向重力性变弱,侧根出现沿水平方向生长的倾向。
【研究背景】
向重力性是植物通过感测重力方向配置身体各器官的重要环境响应特性之一,比如使根部朝着水分和营养成分丰富的地下生长,茎部朝着有利于进行光合作用和繁殖的上方生长。
在向性中,器官弯曲的机制是通过植物激素生长素在器官内不均匀分布控制的。研究认为,根部向重力方向生长时,从地上方向通过中心部传输过来的生长素会均匀分布在根冠(图1上左);当根部倾斜,重力方向改变时,生长素就会更多地分布在重力侧(图1上右)。生长素浓度升高会抑制根部的细胞伸长,因此伸长区域的根部会弯曲(图1下)。这种器官内的生长素分配由生长素外输载体PIN蛋白在细胞内的不均配置控制。
在根和茎等器官中都有感受重力方向变化(器官倾斜)的重力感受细胞。该细胞中含有的质体会大量积聚淀粉形成淀粉体,其比重比细胞质高,会向重力方向沉降。已知这对识别重力方向至关重要。根部的重力感受细胞是位于根冠内的柱状细胞(图1下)。柱状细胞的作用被认为是感受重力方向的变化并向重力侧传输更多的生长素。不过,此前一直不清楚重力感受细胞内的重力信号传递机制。研究团队此前利用模式植物拟南芥确认,植物特有的基因LZY(LAZY1-LIKE)家族在重力感受细胞内参与了重力刺激下生长素的分布调控,但并不清楚其在重力感受细胞内具体负责什么反应。
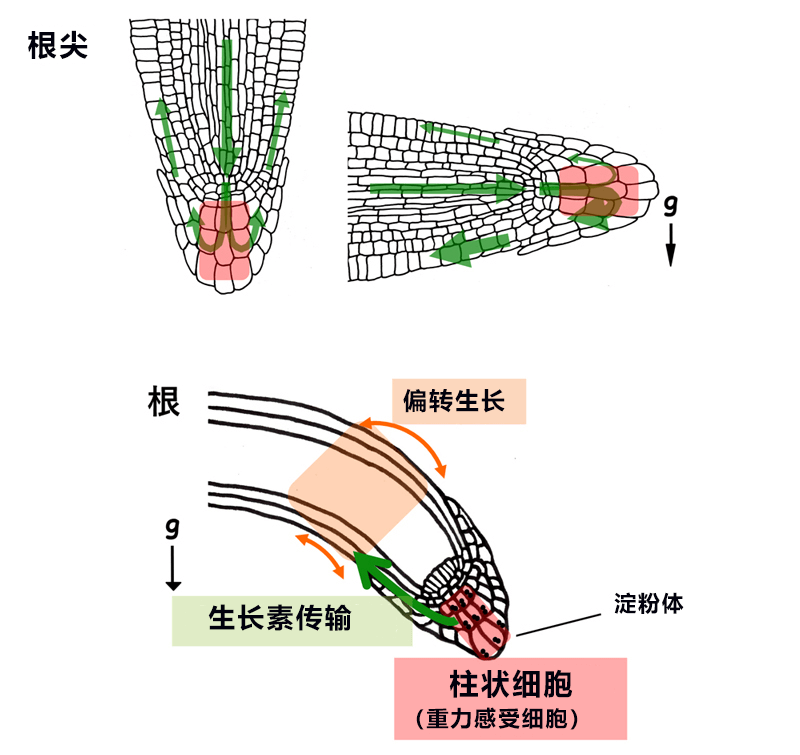
图1:拟南芥根部的向重力性反应
上:柱状细胞(重力感受细胞)在根尖的位置(红色)以及生长素的流动(绿色箭头)。下:从重力感受细胞到器官偏转生长的信息动向。
【研究成果】
研究团队首先确定了与LZY蛋白质相互作用的因子,将具有类似结构的蛋白质命名为RLD(RCC1-like domain protein)1~4,并解析了其功能。解析发现:RLD1和RLD4在根部的重力感受细胞中发挥作用;rld1 rld4双重突变体的向重力性较弱,侧根的生长接近水平方向;对重力进行响应的生长素传输出现异常(图2)。由此可知,RLD与LZY在同一细胞中发挥同样的作用。另外,利用rld1 rld2 rld3 rld4四重突变体进行解析确认,RLD可通过控制PIN蛋白来调节生长素传输。

图2:拟南芥侧根的生长方向
与野生型相比,rld1 rld4双重突变体和lzy3突变体的侧根倾向于水平生长。
研究确认,LZY与RLD是通过位于各自C末端的CCL域与BRX域相互作用。这种结合起着重要作用,决定了这些蛋白质存在于细胞内的哪个位置。在培养细胞中,LZY存在于细胞膜,RLD存在于细胞质,二者分别存在于不同的位置。但当LZY和RLD在同一个细胞中表达时,RLD与LZY都存在于细胞膜中(图3)。另外,二者共同存在于细胞膜依赖于CCL域与BRX域之间的相互作用。此外,研究团队纯化得到了高纯度的CCL-BRX蛋白质复合体,并在1000多种结晶条件下进行筛选后获得了晶体(图4左)。利用大型同步辐射设施(SPring-8,光子工厂)对蛋白质晶体实施了X射线结构解析,成功确定了复合体的三维结构。BRX域的结构为首次报道。BRX域呈简洁紧凑的形状,能识别CCL独具特色的β发夹结构。另外,还在原子水平上明确了BRX与CCL通过形成分子间β片层而强力结合的机制(图4右)。研究团队根据该结构信息,向LZY或RLD导入预计会抑制相互作用的点突变。结果发现,两种蛋白质之间的相互作用消失,而且即使存在LZY,RLD也只停留在细胞质中。另外,二者在重力感受细胞中的功能也丧失。这表明,LZY将RLD引入细胞膜的性质对其在重力感受细胞中的功能很重要。
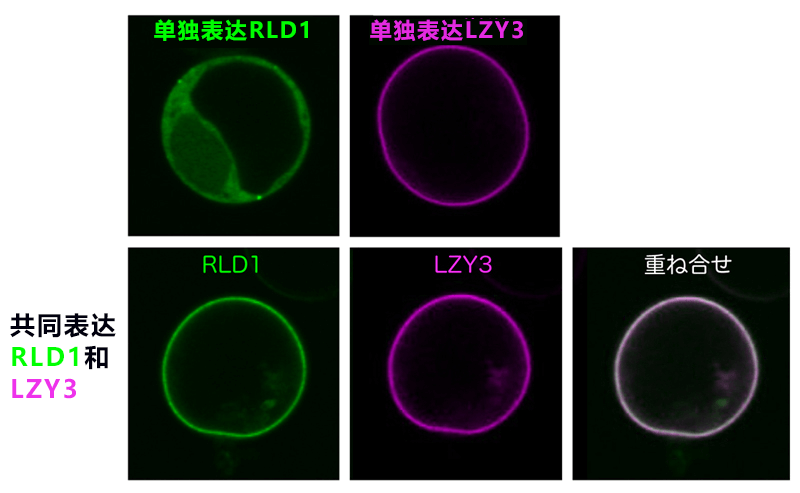
图3:RLD通过LZY存在于细胞膜中
上:使培养细胞原生质体化,分别单独表达RLD1和LZY3。RLD1存在于细胞质,LZY3存在于细胞膜中。下:同时表达RLD1和LZY3后,RLD1与LZY3一起存在于细胞膜中。
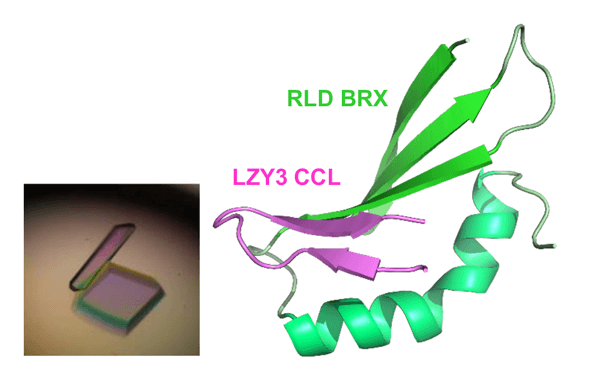
图4. 蛋白复合体结构
左:LZY CCL域与RLD BRX域的蛋白复合体晶体
右:CCL-BRX复合体的X射线晶体结构
研究团队分析了LZY和RLD在重力感受细胞内的工作机制。植物中的LZY蛋白质含量非常低,因此极难观察。通过仔细挑选用于观察的转化子,并使固定的侧根透明化,最终在作为重力感受细胞的柱状细胞中成功捕获了微量的LZY蛋白质。有趣的是,LZY集中存在于重力侧的细胞膜;旋转180度并进行重力刺激30分钟后,其出现在了新的重力侧的细胞膜中(图5)。此时,淀粉体也已经沉降到新的重力侧。RLD通常在柱状细胞内几乎只出现于细胞质中;同样实施重力刺激60分钟后,RLD也出现在了新的重力侧的细胞膜中。在lzy突变体中则未观察到这种现象。不过,LZY集中存在于重力侧的现象在rld突变体中也被观察到。另外,研究团队调查在柱状细胞中发挥作用的PIN3蛋白在根冠中的分布情况时发现,在实施重力刺激5小时后野生型集中分布于重力侧,而rld1 rld4突变体中这种集中分布大为减少。此外,在重力感受细胞中的人为过量表达LZY时,LZY会均匀分布于整个细胞膜中;在这种情况下,RLD也均匀分布在整个细胞膜中。此时,侧根的生长方向会分散,由此可见,适度调节LZY蛋白质的量,并使其保持偏向于重力侧的分布对向重力性至关重要。
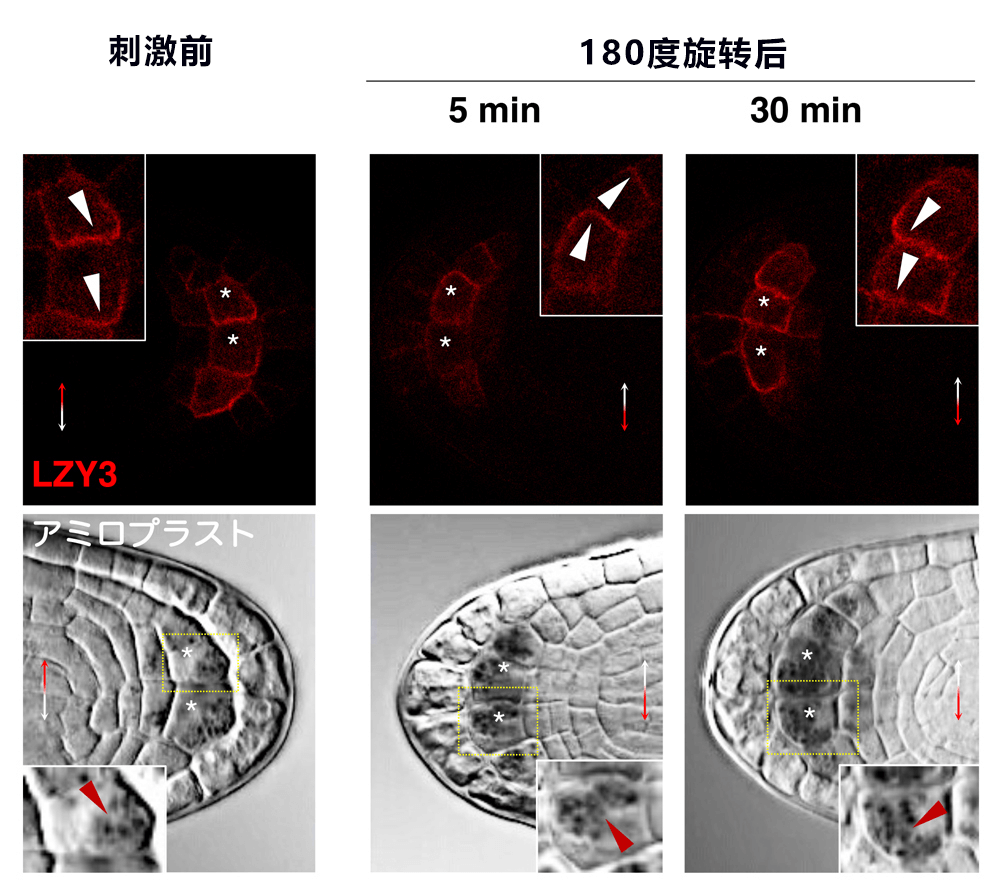
图5:LZY3存在于重力方向侧的细胞膜中
上:固定幼侧根并使其透明化后观察了LZY3。实施刺激前,LZY3集中分布在重力侧的细胞膜中。旋转180度并实施重力刺激,5分钟后,LZY3仍位于原来的重力方向,但30分钟后则集中分布在新的重力方向。下:淀粉体在5分钟后几乎没移动,但30分钟后移动到了新的重力方向。*:中央柱状细胞,白色箭头:LZY3的不均匀分布,红色箭头:淀粉体。
研究团队根据以上结果提出了下述重力信号传递机制:在侧根柱状细胞中,①LZY基于淀粉体的位置信息集中分布在重力侧的细胞膜中;②RLD根据CCL-BRX间的相互作用从细胞质向重力侧的细胞膜移动;③基于RLD的控制会使PIN蛋白向重力侧集中;④更多地向重力侧传输生长素(图6)。
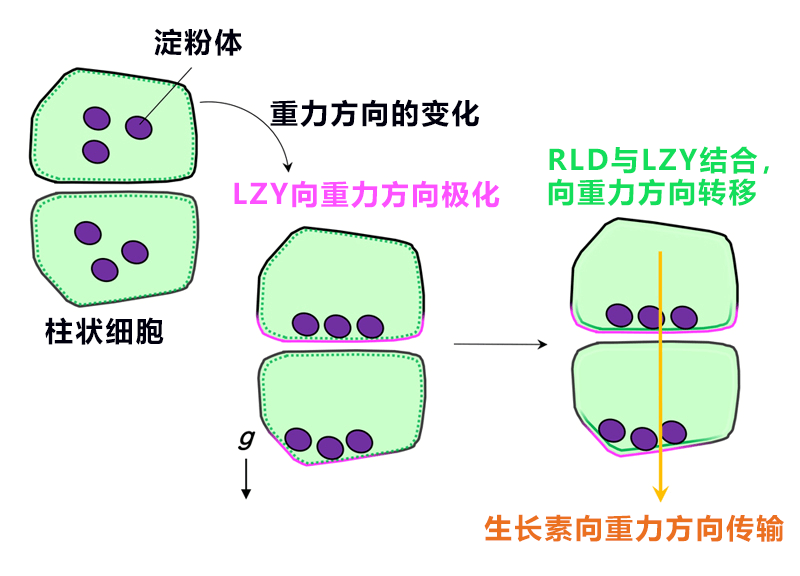
图6:侧根柱状细胞的重力信号传递
【未来展望】
向重力性还参与侧枝和侧根的生长角度控制,是通过植物器官的空间配置来控制株型(即枝条构型、根系构型)的主导因素之一,是育种的重要性状。虽然在农业上属于重要性状,但始终不太了解其分子机制。此次,通过解析RLD的表达与LZY之间的功能性关系,具体明确了部分分子机制,比如淀粉体沉降后如何向重力侧传递信息。本次研究取得的成果将大大促进对向重力性中的重力信号传递分子机制的理解。研究团队的森田教授表示:“今后打算进一步调查LZY集中存在于重力侧细胞膜中的机制,以查清植物的重力方向识别机制。另外,通过进一步详细解析RLD对PIN的调节,希望能了解对植物至关重要的生长素的传输调控机制”。
论文信息
题目:Polar recruitment of RLD by LAZY1-like protein during gravity signaling in root branch angle control
期刊:《Nature Communications》
DOI:https://doi.org/10.1038/s41467-019-13729-7
文:JST客观日本编辑部翻译编译